Jian-wei Liu1,
Chong-wei Hao2,
Zhao-hui Wang1,
Dan Jin3 ![]()
For correspondence:- Dan Jin Email: jdjindan43@hotmail.com Tel:+862061641114
Received: 9 October 2015 Accepted: 5 February 2016 Published: 31 March 2016
Citation:
Liu J, Hao C, Wang Z, Jin D.
Nobiletin inhibits ex
© 2016 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To investigate the anti-inflammatory effects of nobiletin on human osteoarthritic chondrocytes and also to explore possible related molecular events.
Methods: Isolated human osteoarthritic chondrocytes were stimulated with IL-1β. The effect of nobiletin (75, 150 or 300 µg) on chondrocyte viability was assessed. Furthermore, the effect on NO production was determined using Griess reagent while the levels of IL-6 and PGE2 were assessed by enzyme linked immunosorbent assay (ELISA). The influence of nobiletin on the ex
Results: Nobiletin (75, 150 and 300 µg) significantly (p < 0.05) improved the viability of chondrocytes, and remarkably reduced the levels of NO, IL-6 and PGE2. The ex
Conclusion: Inflammatory cytokines are critically involved in the pathogenesis of OA. Significant suppression of cytokines and modulation of PI3/Akt and MAPK signalling cascades by nobiletin suggests its potent anti-inflammatory and anti-osteoarthritic effects.
Introduction
Osteoarthritis (OA) is a degenerative musculoskeletal disease characterized by progressive damage and erosion of articular cartilage, osteophyte formation and subchondral bone sclerosis eventually leading to pain and disability [1,2]. Chondrocytes of the articular cartilage play critical roles in pathogenesis of OA by potentially expressing and as well responding to inflammatory cytokines [3]. These inflammatory cytokines perturb the homeostasis of metabolic processes in the joints [4].
IL-1β, a major cytokine in OA [5] induces the expression of matrix metalloproteinases (MMPs) such as collagenases (MMP-3 and -13). MMPs are involved in cartilage degradation that eventually leads to the release of matrix proteoglycans, a critical event in OA pathogenesis [6]. IL-1β induces activation of NF-κB and MAPK signalling pathways in chondrocytes thereby enhancing the expression and release of NO and PGE2 via up-regulation of cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) [7]. Studies have shown that NO up-regulates the production of MMPs and inflammatory cytokines in OA [8]. Moreover, studies have suggested activated phosphatidylinositol 3-kinase (PI3K) - protein kinase B (Akt) pathway to be associated in regulating chondrocyte survival [9]. Thus compounds that could inhibit the effects of IL-1β and modulate the NF-κB, PI3K/Akt and MAPK signalling pathways and prevent cartilage degradation are of great clinical value in the therapy of OA.
Recently, several researchers have focused on the effects of plant-derived compounds in curbing inflammatory responses in arthritis. Astragalin, a bioactive component from Rosa agrestis [10] and piperine derived from Piper nigrum exhibited anti-inflammatory effects in OA model [11]. In our study, we investigated the effects of nobiletin in IL-1β-induced OA. Nobiletin, a polymethoxylated flavone found in citrus peels has been reported to possess various biological properties as anti-cancer [12], anti-proliferative [13], anti-inflammatory effects [14]. The present study was taken as an effort to explore the molecular events associated with the anti-inflammatory effects of nobiletin.
Methods
Chemicals and reagents
Nobiletin was purchased from Sigma-Aldrich (St. Louis, MO, USA). Antibodies against p38, JNK, ERK, p-p38, p-ERK, p-JNK, p-NF-κB p65, p-IκBα (Cell Signalling Technology Inc. Beverly, MA) Bad, Bax, Bcl-2, Bcl-xL, Akt, p-Akt, GSK-3β, p-GSK-3β, mTOR, PTEN, iNOS, TNF-α and COX-2 (Santa Cruz, CA,USA) were used for western blotting. Griess Reagent was purchased from Beyotime Institute of Biotechnology (Shanghai, China). DMEM, FBS, collagenase II, penicillin and streptomycin were purchased from Gibco BRL (Grand Island, NY, USA). Recombinant human IL-1β was purchased from (R&D systems. Minneapolis, MN, USA). All other chemicals used in the study were of analytical grade and were procured from Sigma-Aldrich (St. Loius, MO, USA) otherwise are specified.
Chondrocyte isolation and cell culture
Articular cartilage samples from 25 informed and consented patients (age: 57 ± 13) who were undergoing total knee replacement surgery were obtained. The study was approved by the institutional ethical committee. The articular cartilage was harvested from non-lesional areas and primary chondrocytes were isolated as described by Cheng et al [15].
In brief, the tissues were digested with trypsin (0.25 %) for 30 min and further digested using collagenase II (0.2 %) in DMEM for 6 h at 37 °C. The cells were suspended in DMEM with 10 % fetal bovine serum (FBS), 100 U/mL of penicillin and 100 mg/mL of streptomycin and cultured at 37 °C with 5 % CO2. On reaching 80 % confluence, the cells were pre-treated with nobiletin at 70, 150 or 300 μg/mL for 6 h and were subsequently treated with IL-1β (5 ng/mL) for 24 h.
Cell viability assay
The effect of nobiletin on the viability of chondrocytes was assessed by the MTT assay. The cells were seeded at a density of 6 x 103 in a 96-well plate and were cultured overnight, following which the cells were treated with various concentrations of nobiletin for 24 h. The medium was discarded and 20 μL MTT (5 mg/mL) was added to each well and incubated for 4 h. After discarding the supernatant, 150 μL of DMSO was added to solubilize the formazan crystals. The absorbance was measured at 570 nm using a micro-plate reader (Bio-Rad, Hercules, CA, USA).
Annexin V/PI staining
Cell apoptosis was measured by annexin V-FITC and PI staining. Chondrocyte cells that were exposed to nobiletin for 6 h and IL-1β for 24 h were seeded in a 24-well plate at a density of 2 × 105 cells per well. Apoptosis was detected using apoptosis detection kit (Santa Cruz Biotechnology, Santa Cruz, CA, USA). The cells were washed in PBS (phosphate buffered saline) and were then resuspended in binding buffer containing 100 μL FITC-conjugated anti-annexin V antibodies. The cells were analyzed for fluorescence using a flow cytometer (FACS Calibur, BD Biosciences).
Determination of IL-6 and PGE2
Chondrocytes were pre-treated with nobiletin for 3 h and then stimulated with IL-1β for 24 h. The concentrations of IL-6 and PGE2 in the culture medium were measured by ELISA (R & D Systems, Minneapoils, MN).
NO assay
NO concentration in the culture medium of the chondrocytes treated with various concentrations of nobiletin and IL-1β as described above for IL-6 determination was assessed using Griess reagent as previously described by Au et al [16].
RNA isolation and RT-PCR
Real-time PCR was used to detect the expression of iNOS and COX-2. Total RNA was extracted using TriZol (Invitrogen) according to the manufacturer's instructions and quantified. RNA was quantified spectrophotometrically at 260 nm (HP 8452A Diode Array Spectrophotometer). First strand cDNA was synthesized using 0.3 mg of the isolated RNA using DyNamoTM cDNA Synthesis Kit (Fermentas). Synthesized cDNA were amplified by PCR using the following primers: iNOS Forward - CCTTACGAGGCGAAGAAGGACAG, Reverse - CAGTTTGAGAGAGGAGGCTCCG; COX-2 Forward - GAGAGATGTATCCTCCCACA GTCA, Reverse – GACCAGGCACCAGACCA AAG. The relative expression of COX-2 and iNOS were normalized with GADPH (primers - Forward - TCTCCTCTGACTTCAACAGCGAC, Reverse - CCCTGTTGCTGTAGCCAAATT C). PCR was carried out as described by Ying et al [11].
Western blot analysis
Proteins extracted from chondrocytes treated with nobiletin (70, 150 or 300 µg/mL) and stimulated with IL-1β were subjected to western blotting. The cells were subjected to lysis buffer and the nuclear and cytoplasmic proteins were extracted from the cells using NE-PER Mammalian Protein Extraction Reagent (Thermo). Concentration of the isolated protein was determined using BCA protein assay kit (Bio-Rad Laboratories, USA). Equal amounts of protein samples (40 μg) were separated on SDS-polyacrylamide gel (12 %) and transferred to PVDF membranes. The membranes were blocked with 5 % non-fat dry milk and were incubated with primary antibodies overnight at 4 °C followed by incubation with peroxidase-conjugated secondary antibodies at room temperature for 60 min. The immunoreactive bands were detected by enhanced chemiluminescence (GE Healthcare).
Statistical analysis
Data are presented as mean ± SD (n = 3 or 6). Statistical significance between the means of various experimental groups were assessed by one-way analysis of variance (ANOVA) followed by Duncan’s Multiple Range Test as post-hoc analysis. Statistical difference at p < 0.05 was considered significant.
Results
Cytotoxic effects of nobiletin
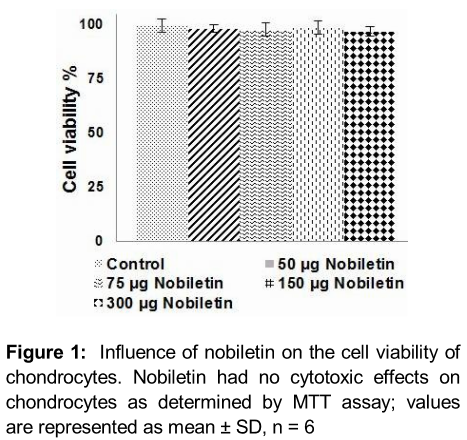
Articular chondrocytes incubated with nobiletin (75, 150 or 300 µg/mL) were subjected to MTT assay to assess if nobiletin had any cytotoxic effects. We observed that at all tested concentrations there was negligible effects on viability of chondrocytes (). The results suggest that nobilein exhibited no cytotoxic effects.
Nobiletin improves viability of chondrocytes exposed to IL-1β
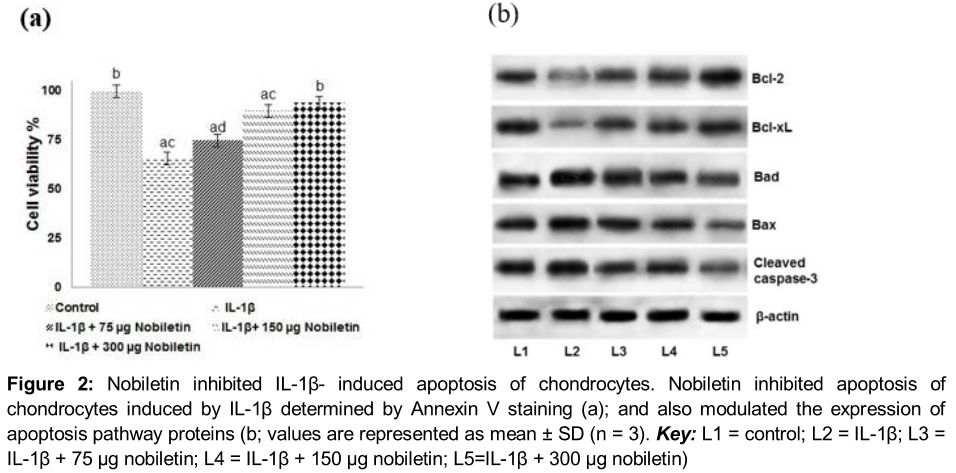
Annexin V/PI staining and flow cytometry assay was employed to assess apoptosis of chondrocyte on exposure to IL-1β. We observed apoptosis of chondrocytes stimulated with IL-1β. However, pre-treatment with nobiletin interestingly caused a decrease (p < 0.05) in apoptosis, with 300 µg concentration of nobiletin exhibiting maximum protective effects (). The low dose of 75 µg though decreased apoptosis, the trend of reduction was significantly lesser as compared to higher concentrations of 150 and 300 µg. The results indicated that nobiletin effectively inhibited IL-1β- toxicity of chondrocytes.
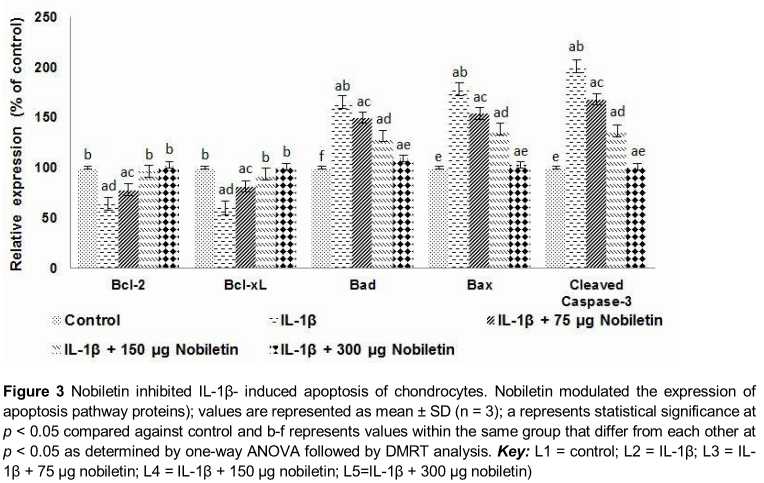
Further to assess the influence of nobiletin on the expression of apoptotic pathway proteins, we analysed the expression of pro-apoptotic (Bax and Bad) and anti-apoptotic (Bcl-2 and Bcl-xL) proteins. IL-1β-stimulated chondrocytes expressed enhanced levels of pro-apoptotic proteins with decreased expression of Bcl-2 and Bcl-xL (), suggesting raised apoptotic counts observed could have been due to raised levels of apoptotic protein expression. Interestingly, nobiletin caused significant up-regulation in the levels of Bcl-2 and Bcl-xL while it down-regulated the expression of Bax and Bad. Thus, the results suggest the anti-apoptotic effects of nobiletin.
Nobiletin inhibits IL-1β-induced NO and PGE2 production
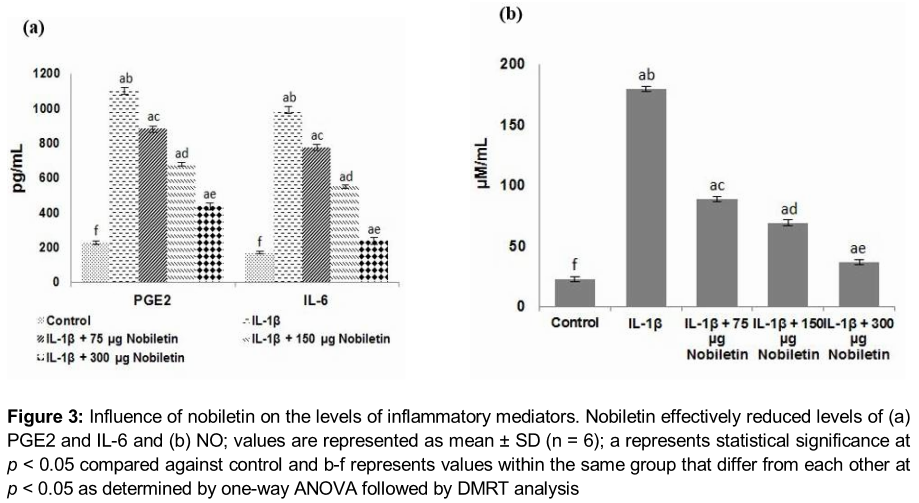
NO and PGE2 levels in the culture medium of chondrocytes stimulated by IL-1β were determined. Markedly (p < 0.05) enhanced levels of NO and PGE2 were observed. Multi-fold increases in the level of NO were seen. Moreover, increased levels of inflammatory cytokine, IL-6 were also observed upon IL-1β exposure (). Nobiletin caused significant (p < 0.05) decrease in the levels of NO, PGE2 and 1L-6. The results suggest the potent anti-inflammatory effects of nobiletin. The 150 and 300 µg concentrations were more effective than 75 µg nobiletin.
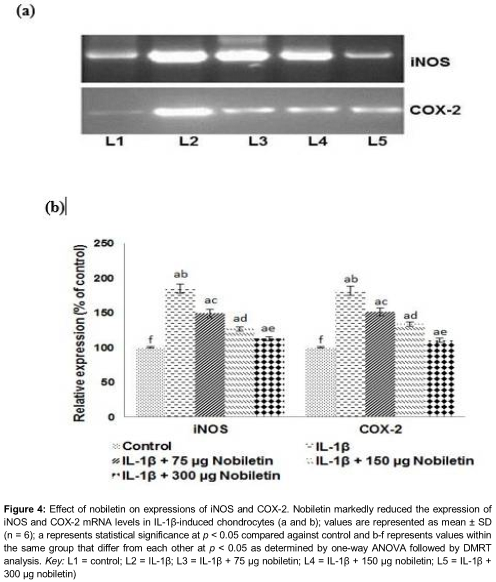
Nobiletin suppressed iNOS and COX-2 expression
IL-1β is known to induce the expression of COX-2 and iNOS [7], eventually leading to the production of PGE2 and NO. Here we investigated whether nobiletin influenced expression of iNOS and COX-2. In line with previous studies, we also observed markedly elevated mRNA iNOS and COX-2 expression () and as well raised expression of iNOS and COX-2 at the protein levels () in chondrocytes stimulated with IL-1β. Nobiletin pre-treatment for 6 h caused a multi-fold decrease in the expression levels of both COX-2 and iNOS. The reductions in the expression at both the mRNA and protein levels were found to be dose-dependent, with 300 µg concentration exerting maximum level of reduction. The reduction in the expression of IL-6, NO and PGE2 were in line with down-regulated COX-2 and iNOS. The observations reveal that nobiletin-induced decreased mRNA levels which could have contributed to reduced expressions of IL-6, NO and PGE2.
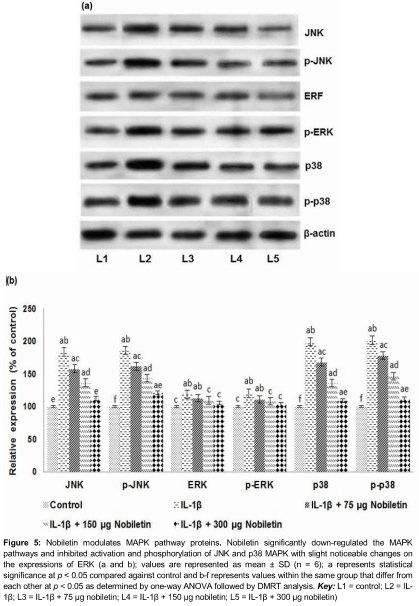
Effect of nobiletin on expression of MAPK cascade proteins
We assessed the expression of phosphorylated JNK, ERK and p38 MAPK. IL-1β stimulation caused a striking increase in the expressions of activated JNK, ERK and p38 MAPK (). Previous studies have shown enhanced expression of MAPKs under IL-1β stimulation [17]. However, nobiletin exposure resulted in significant down-regulation in the activation level of JNK, ERK and p38 MAPK. The phosphorylated JNK and p38 MAPK were highly reduced as compared to the p-ERK. The effects were similar at all the concentrations of nobiletin. The expression of ERK though reduced on exposure to nobiletin, the reduction was not significant as compared to JNK or p38MAPK.
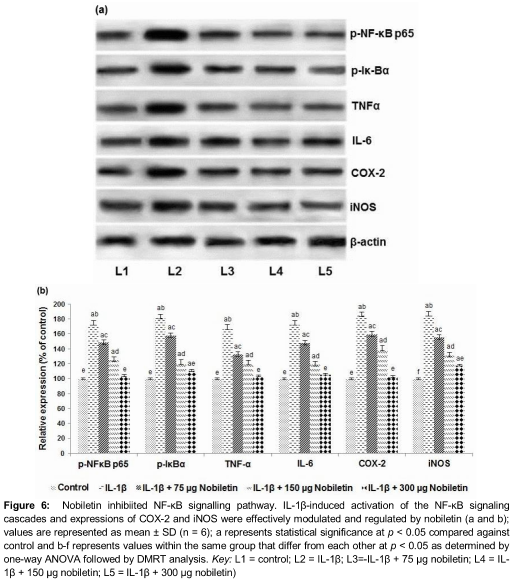
Nobiletin down-regulates NF-κB signalling
NF-κB is a transcription factor that critically regulates expression of inflammatory mediators. To investigate whether nobiletin exhibited anti-inflammatory effects via modulating the NF-κB pathway, the expression of NF-κB pathway proteins were detected. IL-1β caused significantly up-regulated expression of p-NF-κB p65, p-IκBα, IκBα and TNF-α. Pre-treatment with nobiletin for 6 h markedly (p < 0.05) down-regulated the expression in a dose-dependent manner (). This down-regulation of the pathway could have possibly contributed to the decreased expression of inflammatory mediators.
Influence of nobiletin on the PI3K/Akt signalling pathway
To investigate the anti-inflammatory mechanisms of nobiletin, we also assessed the expression of Akt pathway proteins. IL-1β-induced chondrocytes exhibited elevated levels of Akt, phosphorylated Akt, phosphorylated GSK-3β and mTOR with decreased expression levels of PTEN. Nobiletin at all the three analysed concentration was observed to significantly (p < 0.05) inhibit the phosphorylation of Akt and GSK-3β (). Furthermore, PTEN expression was found to be up-regulated by nobiletin in a dose-dependent manner. Nobiletin thus effectively regulates the PI3K/Akt signalling pathway and this in part also contributed to the decreased expressions of NF-κB.
Discussion
Osteoarthritis (OA) is a degenerative disease of the synovial joints that reduces quality of life [18]. Anti-inflammatory drugs prescribed for treating osteoarthritis are generally recommended for long term use. As individuals respond differently to drugs, chances of potential side effects remain as the major problem due to long term use of anti-inflammatory drugs. Recently, use of phytochemicals as anti-inflammatory agents is considered as an effective strategy to block inflammation in many inflammatory diseases such as OA [19]. In our study, we investigated the anti-inflammatory effects of nobiletin on IL-1β-induced inflammation in human osteoarthritic chondrocytes.
Role of inflammatory mediators NO and PGE2 is well known in the pathogenesis of OA and increased production of NO and PGE2 have been reported in patients with OA [20]. IL-1β, an important inflammatory cytokine, is a major catabolic factor associated with OA exerting critical roles in cartilage degradation [21]. Elevated levels of IL-1β have been observed in synovial fluid in OA [22]. IL-1β induces the expression of MMPs, such as MMP1, MMP3 and MMP13 that contribute to matrix degradation [23]. In addition, IL-1β induces the expression of iNOS and COX-2 and as well enhance the production of NO, PGE2 and IL-6 [11].
In the present study, we observed significant decrease in the levels of IL-6, NO and PGE2 as well as the expression of iNOS and COX-2 both at the mRNA and protein levels in response to pre-treatment with nobiletin. These observations suggest that nobiletin was able to effectively reduce the expression of inflammatory mediators, a hallmark event in OA therapy. Nobiletin improved chondrocyte cell viability in a dose-dependent manner and had no cytotoxic effects on chondrocytes that were not exposed to IL-1β. Nobiletin drastically altered the expression of apoptotic pathway proteins. Up-regulation of anti-apoptotic proteins with significant inhibition of pro-apoptotic proteins and caspase-3 in a dose-dependent way were also observed.
It is known that NF-κB and MAPK signalling pathways play critical roles in the regulation of inflammatory mediators. To further analyse the effects of nobiletin, we assessed the expression of NF κB and MAPK signalling pathway proteins. NF-κB is one of the most important cytokine-induced transcription factors that critically regulate the expression of MMPs, COX-2, iNOS and other inflammatory cytokines [23]. Normally, NF-κB is bound to IκB inhibitor proteins in the cytoplasm. IL-1β stimulation induces p65 phosphorylation and subsequent degradation of IκB that eventually leads to translocation of NF-κB p65 into the nucleus that causes up-regulation of inflammatory mediators [24]. In our study, nobiletin treatment caused remarkable down-regulation of phosphorylated NF-κB p65 and IκBα. The observed decreased expression of COX-2, iNOS and TNF-α suggest the inhibition of NF-κB signalling pathway by nobiletin that possibly could have also contributed to decreased levels of PGE2 and NO.
Previous studies have shown increased expression of MMPs, COX-2, and iNOS following stimulation of chondrocytes by IL-1β [25]. Thus, the results indicate that nobiletin exerts anti-inflammatory effects via attenuation of NF-κB activity. Further, it has been reported that NF-kB signalling pathway mediates many of the biological effects of IL-1β on chondrocytes [26]. Thus inhibition of the pathway could be of immense clinical value in treatment of OA.
PI3K and Akt are the upstream molecules in NF-κB pathway and have critical role in NF-κB activation [18]. To assess the effects of nobiletin on PI3K/Akt signalling, the expression of the pathway proteins was analysed. IL-1β caused multi-fold increased expression of phosphorylated Akt and GSK-3β, indicating activation of the pathway. Nobiletin pre-treatment significantly inhibited the pathway as observed by decreased phosphorylation of Akt. The results suggest the possible involvement of PI3K/Akt cascades in nobiletin-induced inhibition of NF-κB activation.
MAPK pathways also exert critical effect the regulation of inflammatory cytokines production [27]. Binion et al. [28] reported that the activation of Akt further caused activation of MAPK pathways. Stimulating chondrocytes by IL-1β has been shown to induce phosphorylation and activation of ERK1/2, JNK, and p38 MAPKs [17]. In our study, enhanced expression of MAPK pathway proteins JNK, ERK and p38 were observed following IL-1β which is similar to the results of previous studies [10, 17]. Nobiletin down-regulated the MAPK activation as evidenced by significant decrease in the expression of JNK, ERK and p38. Comparatively with the extent of JNK and p38 MAPK expression, ERK expression did not exhibit drastic changes, though some noticeable changes were seen. Nobiletin was able to effectively down-regulate MAPK pathways in IL-1β stimulated chondrocytes which also in part could be responsible for reduced levels of inflammatory mediators observed.
Conclusion
Collectively, the observed results reveal the ant-inflammatory effects of nobiletin. Since nobiletin effectively modulates the critical pathways involved in OA pathogenesis, it requires further investigation as a potent candidate in OA therapy.
References
Archives


News Updates